Neutralizing monoclonal antibodies from convalescents can prevent SARS-CoV-2 infection in vivo
To obtain neutralizing mAbs from convalescents, we collected peripheral blood mononuclear cells (PBMCs) from the two earliest Wuhan COVID-19 convalescents at one year after SARS-CoV-2 WT infection. PBMCs were stained with Biotin-labeled S1 protein to sort antigen-specific memory B cells (MBCs), and a total of 226 S1+ single MBCs were successfully sorted into 96-well plates with frequencies of 0.70%. After screening 113 successfully expressed antibodies by SARS-CoV-2 pseudovirus neutralization assay, six potent neutralizing antibodies (3C4, 4G4, 7B3, 12G4, 13B2, and 14B1) were selected for further investigation.
The enzyme-linked immunosorbent assay (ELISA) results showed that all six mAbs were able to bind the RBD of SARS-CoV-2 Spike (Fig. 1a), even though S1 was used as the bite in MBCs sorting. We speculated that the antibodies that bound to RBD could more efficiently block the interaction between Spike and hACE2 and outcompeted in the initial screen of pseudovirus neutralization. Meanwhile, the 50% effective concentration (EC50) of these mAbs against SARS-CoV-2 pseudovirus (carrying the S of the SARS-CoV-2 WT strain with D614G mutation, WT-D614G) was determined, and three mAbs showed potent neutralizing activity. The EC50 values of 4G4, 7B3, and 14B1 were 3.7 ng/mL, 1.8 ng/mL, and 3.5 ng/mL, respectively (Fig. 1b).

Monoclonal antibodies 7B3 and 14B1 can neutralize SARS-CoV-2 in vitro and in vivo. a The RBD binding assay of mAbs by ELISA. b The pseudovirus neutralization assay of mAbs against SARS-CoV-2 WT-D614G. c Time schedule for antibody protection in vivo. 12–18-week-old K18-hACE2 mice (n = 5 for each group) were injected 10 mg/kg antibodies at dpi −1, and were intranasally infected with 250 PFU/mL SARS-CoV-2 WT strain viruses each at dpi 0. Endpoint is at dpi 8 or when mice dead or reach 80% body weight, and the mice were euthanized and samples at endpoint. d Body weight changes. e Survival rate. f The SARS-CoV-2 RNA (E and N gene) copy number in lungs and brains. Unpaired t test. *p < 0.05; **p < 0.01; ****p < 0.0001. g The H&E stain of lungs. Scar bar, 200 μm in the up row, 50 μm in the down row. h The immunofluorescence analysis of SARS-CoV-2 N protein in the lungs. Nuclear DNA is stained by DAPI (4’,6-diamidino-2-phenylindole). Scar bar, 50 μm
Next, we evaluated the prophylactic effects of 4G4, 7B3, and 14B1 against SARS-CoV-2 infection in K18-hACE2 knock-in (KI) mice, which are highly susceptive to SARS-CoV-2. 12 ~ 18-week-old K18-hACE2 KI mice were intraperitoneally (i.p.) injected with 10 mg/kg mAbs on day −1 and then were infected intranasally with 250 PFU/mL SARS-CoV-2 WT viruses at day 0. REGN10933, which recognizes RBD and is an effective component of FDA-proved mAbs cocktail (REGN-COV2), was included as the positive control. The mice treated with the isotype control dramatically lost body weight from 3 dpi, and all succumbed to infection by 8 dpi. However, mice treated with 4G4, 7B3, or 14B1 all survived, and no significant weight change was observed (Fig. 1c–e). The viral loads of SARS-CoV-2 in the lungs and brains of K18-hACE2 KI mice were also determined. 4G4, 7B3, and 14B1 significantly decreased the viral loads in the lungs and brains compared to the isotype control (isotype) mAb (Fig. 1f). Lung pathology analysis showed that SARS-CoV-2 caused severe interstitial pneumonia characterized by a large number of inflammatory cells infiltrating the lungs and the wide thickening and rupture of the alveolar septum in the isotype group. In contrast, no obvious lesions were observed in mice injected with 4G4, 7B3, or 14B1 (Fig. 1g). Meanwhile, a large amount of SARS-CoV-2 antigen (Nucleocapsid, N protein, green) was detected in the lungs of mice treated with the isotype control, but little N protein was observed in mice treated with our mAbs (Fig. 1h). These results indicated that 4G4, 7B3, and 14B1 can efficiently protected the K18-hACE2 KI mice from SARS-CoV-2 WT infection.
Epitope competition analyses
We next quantitatively measured the binding capacity of mAbs for SARS-CoV-2 WT-RBD through biolayer interferometry (BLI). The kinetic binding affinity (KD) values of 7B3 and 14B1 were 2.6 × 10–12 mol/L and 3.1 × 10–12 mol/L, respectively, consistent with their high neutralizing activity against WT-D614G pseudovirus (Table 1). Receptor blockage is reported as the primary strategy for neutralizing antibodies to prevent SARS-CoV-2 infection. We further investigated the competition capacity of our mAbs and hACE2 for RBD binding by BLI. This analysis showed that the six mAbs recognized three major epitopes on RBD. Except for 13B2, all five mAbs blocked the binding of hACE2 to RBD (Supplementary Fig. 1a), partly explaining the inefficient neutralizing activity of 13B2 against SARS-CoV-2. 12G4 competes with hACE2, but the binding of 12G4 to RBD did not affect the binding of RBD by the other five mAbs. The highly potent neutralizing mAb 14B1 partially competed with 4G4 and 7B3, but severely blocked the interaction of RBD with 3C4. Taking the neutralizing activity into consideration, we propose that the combination of 7B3 and 14B1 has the potential to be used as antibody cocktails (Fig. 2 and Supplementary Fig. 1b).
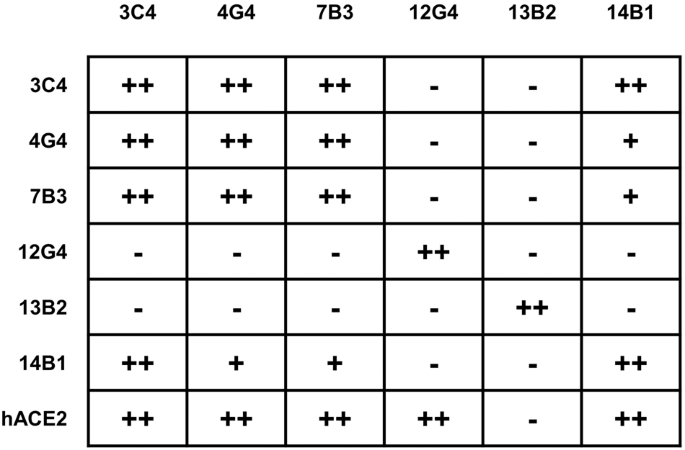
The epitope competition between mAbs. The mAbs in row were loaded onto protein A biosensors, and WT-RBD was then associated. The second mAbs or hACE2 were then loaded to detect the competition relationships (details in Supplementary Fig. 1). − no competition, + partial competition, ++ strong competition
Cross-variant neutralization against major SARS-CoV-2 variants
SARS-CoV-2 has continued to mutate since its outbreak at the end of 2019, and there are five variants of concern so far, namely, Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2), and Omicron (B.1.1.529, BA.1, BA.2, BA.3, BA.4, BA.5, XBB, BA.2.75, BA.5.2, BF.7, XBB.1.5, XBB.1.16, and XBB.2.3.2). The amino acid alterations in the S protein of the variants are summarized in Supplementary Fig. 8. We further evaluate the binding capacity of mAbs against RBD of SARS-CoV-2 Delta, BA.1, and XBB.1.5 through BLI and ELISA, and found out that all six mAbs bind to WT- and- Delta-RBD with varying degrees (Supplementary Fig. 2). Then, the neutralizing activity of these six mAbs against several SARS-CoV-2 variants were investigated, including five VOCs, Deltacron, and Eta by pseudovirus neutralization assay and found that 7B3 potently neutralized the Alpha and Delta variants with EC50 values of 5.118 ng/mL and 18.92 ng/mL, respectively, but 7B3 completely lost its neutralizing activity against the Beta, Gamma, and Eta variants. Meanwhile, 14B1 showed potent and cross-variant neutralizing activity: its EC50 against Alpha, Beta, Gamma, Delta, and Eta variants were 12.79 ng/mL, 30.94 ng/mL, 14.22 ng/mL, 29.11 ng/mL and 23.92 ng/mL, respectively (Fig. 3a and Supplementary Fig. 3).

The neutralizing ability of mAbs against SARS-CoV-2 pseudovirus and authentic virus. a The half effective concentration (EC50) of mAbs against pseudovirus of different SARS-CoV-2 variants. b The authentic SARS-CoV-2 neutralization of mAbs through plaque reduction assay
The Omicron variant, which is currently the dominant variant around the world, has many subvariants, all of which have extensive immune escape ability. We further tested our antibodies against multiple Omicron subvariants: BA.1, BA.2, BA.3, BA.4/5, BA.2.12.1, XBB, BA.2.75, BF.7, BQ.1.1, XBB.1.5, XBB.1.16, and XBB.2.3.2. Encouragingly, 7B3 neutralized BA.1, BA.2, BA.3, and BA.2.75 efficiently with EC50 values of 64.26 ng/mL, 156.6 ng/mL, 34.23 ng/mL and 73.11 ng/mL, respectively. With respect to BA.4/5, BA.2.12.1, BF.7, and BQ.1.1, all six mAbs lost their neutralizing activity (Fig. 3a and Supplementary Fig. 3). Interestingly, 14B1 potently neutralized BA.2.75 with an EC50 value of 41.61 ng/mL, while 14B1 showed no neutralizing activity against the other tested Omicron subvariants. We also evaluated the potential of 7B3 and 14B1 for mAb cocktails and found that the combination of these two mAbs expand the neutralizing breadth against SARS2-CoV-2 variants based on pseudovirus neutralization assay. As these two mAbs partially compete the binding to RBD, we did not pursue the 7B3/14B1 cocktail application further. The authentic SARS-CoV-2 plaque reduction neutralization assay was conducted to corroborate the neutralizing results from the pseudovirus neutralizing assay. Same as VSV pseudovirus results, 7B3 and 14B1 showed potent neutralizing activity against authentic SARS-CoV-2 WT strain and Delta variant and 7B3 also potently inhibits authentic Omicron BA.1 and BA.2 with the EC50 values of 259.3 ng/mL and 237.3 ng/mL, respectively (Fig. 3b). 3C4 was the only mAb to neutralize the emerging XBB (EC50 = 645 ng/mL) and XBB.1.5 (EC50 = 174 ng/mL) variants, even though 3C4 showed no neutralizing activity against all of the other tested Omicron subvariants. This is consistent with our binding data, as only 3C4 displayed a detectable binding signal against XBB-RBD among tested six mAbs, with a kinetically-derived binding affinity of 8.37 nM. However, the binding affinity of the mAbs to the RBDs does not correlate well with the neutralization potency against distinct SARS-COV-2 variants, as 12G4 and 13B2 could bind to Delta-RBD with nanomole to picomole affinities (Supplementary Fig. 2a), respectively, the two mAbs exhibit relatively weaker inhibition activity against Delta strain tested by both pseudovirus and authentic viruses (Fig. 3).
7B3 and 14B1 treat SARS-CoV-2 Delta infection in K18-hACE2 KI mice
The Delta variant, once the dominant lineage, has higher infectiousness than the original SARS-CoV-2 and moderate immune escape ability. Considering the potent neutralizing ability of 7B3 and 14B1 against the Delta variant, we evaluated the therapeutic effects of 7B3 and 14B1 in K18-hACE2 KI mice against the SARS-CoV-2 Delta variant; 12 ~ 18-week-old K18-hACE2 KI mice were intranasally infected with 250 PFU/mL Delta variants. One day later, these mice were i.p. injected with 20 mg/kg mAbs followed by daily monitoring.
As shown in Fig. 4, mice injected with 7B3 or 14B1 survived the infection of the Delta variant, and no significant weight change was observed. However, the isotype control group dramatically lost body weight from 6 dpi, and all mice in this group succumbed to infection by 8 dpi. We also found that 7B3 and 14B1 significantly decreased the viral loads in the lungs and brains compared to the isotype control (Fig. 4a–d). Lung pathology analysis showed that the Delta variant also caused severe interstitial pneumonia characterized by a large number of inflammatory cells infiltrating the lungs and the wide thickening and rupture of the alveolar septum in the isotype group. In contrast, only limited inflammatory cell infiltration was observed in mice injected with 7B3 or 14B1 (Fig. 4e). These results were also consistent with the immunofluorescence analysis, little N protein (green) was detected in the lungs of mice administrated with 7B3 or 14B1 (Fig. 4f).

Mabs7B3 and 14B1 treat infection of SARS-CoV-2 Delta variant in vivo. a Time schedule for antibody therapy in vivo (n = 5 for each group). 12–18-week-old K18-hACE2 mice were intranasally infected with 250 PFU/mL SARS-CoV-2 Delta viruses each at dpi 0, and were injected 20 mg/kg antibodies at dpi 1. Endpoint is at dpi 8 or when mice dead or reach 80% body weight, and the mice were euthanized and samples at endpoint. b Body weight changes. c Survival rate. d The SARS-CoV-2 RNA (E and N gene) copy number in lungs and brains. Unpaired t test. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. e The H&E stain of lungs. Scar bar, 200 μm in the up row, 50 μm in the down row. f The immunofluorescence analysis of SARS-CoV-2 N protein in the lungs. Nuclear DNA is stained by DAPI. Scar bar, 50 μm
7B3 treats SARS-CoV-2 Omicron BA.1 and BA.2 infections in K18-hACE2 KI mice
The Omicron variant, carrying more than 30 mutations in the S protein, has become the dominant lineage instead of Delta since 2022.25 Moreover, Omicron-associated lineages, like BA.1, BA.2, BA.3, BA.4, BA.5, XBB.1.5, XBB.1.16, and XBB.2.3.2, are still emerging. Due to a large number of mutations in the S protein, Omicron has higher infectiousness and stronger immune escape ability than the other SARS-CoV-2 variants, causing many breakthrough infections around the world.
Though Omicron can largely escape from most neutralizing antibodies, 7B3 was able to neutralize multiple Omicron subvariants including BA.1, BA.2, and BA.3, we then investigated the therapeutic effect of 7B3 in K18-hACE2 KI mice against the Omicron (BA.1 and BA.2) variants. 12 ~ 18-week-old K18-hACE2 KI mice were infected with 500 PFU/mL Omicron BA.1 variant or 2500 PFU/mL Omicron BA.2 on day 0, and then these mice were i.p. injected with 20 mg/kg mAbs on day 1. Encouragingly, mice treated with 7B3 survived the infection of the BA.1 and BA.2 variants, and no significant weight change was observed. However, the isotype control group dramatically lost body weight from 4 dpi for BA.1-challenged mice or 5 dpi for BA.2-challenged mice, and all the control mice succumbed to infection by 8 dpi (Fig. 5b, c). We also found that 7B3 significantly decreased the viral loads in the lungs and brains compared to the isotype control (Fig. 5d). Lung pathology analysis showed that both Omicron BA.1 and BA.2 caused severe interstitial pneumonia characterized by a large number of inflammatory cells infiltrating into the lungs and the wide thickening and rupture of the alveolar septum in the isotype group. In contrast, only limited inflammatory cell infiltration and slight thickening were observed in mice injected with 7B3 (Fig. 5e). The immunofluorescence analysis also showed that less N protein (green) was detected in the lungs of mice treated with 7B3 than control mice (Fig. 5f). These results indicated that 7B3 can efficiently prevent infection by the Omicron subvariants BA.1 and BA.2 in K18-hACE2 KI mice.

mAbs 7B3 treat infection of SARS-CoV-2 Omicron BA.1 and BA.2 variants in vivo. a Time schedule for antibody therapy in vivo (n = 5 for each group). 12–18-week-old K18-hACE2 mice were intranasally infected with 500 PFU/mL BA.1 or 2500 PFU/mL BA.2 viruses each at dpi 0, and were injected 20 mg/kg antibodies at dpi 1. Endpoint is at dpi 8 or when mice dead or reach 80% body weight, and the mice were euthanized and samples at endpoint. b Body weight changes. c Survival rate. d The SARS-CoV-2 RNA (E gene and N gene) copy number in lungs and brains. Unpaired t test. *p < 0.05; **p < 0.01; ****p < 0.0001. e The H&E stain of lungs. Scar bar, 200 μm in the up row, 50 μm in the down row. f The immunofluorescence analysis of SARS-CoV-2 N protein in the lungs. Nuclear DNA is stained by DAPI. Scar bar, 50 μm
The Cryo-EM structure of 14B1 in complex with SARS-CoV-2 WT Spike
To understand the 14B1-mediated neutralization mechanism, we obtained cryo-electron microscopy (cryo-EM) reconstruction of 14B1 Fab in complex with the ectodomain of the trimeric spike protein (WT-Spike-6P) at an overall resolution of 3.2 Å. Only one Spike state was identified, with three up-RBDs occupied by three 14B1 Fabs (Fig. 6a). To obtain more detailed information on antibody-antigen interactions, we focused on the RBD-variable fragment (Fv) of 14B1 and generated a local refined map (Supplementary Fig. 4). Structural analysis shows that 14B1 resides on the top of RBD, and the binding site overlaps partially with binding residues made by hACE2 (Fig. 6b), belonging to the RBD-2 antibody in the Hastie’s classification system.19

Cryo-EM reconstruction of 14B1-RBD complex. a (Left) Cryo-EM structure of three 14B1 Fab in complex with SARS-CoV-2 S-6P trimer. All three Fabs (light chain colored in the green, heavy chain in slate blue) bind on the top face of RBD, with one of three spike protomers colored in beige and the other two in gray. The local refinement region is boxed with black box. (Right) Side view of 14B1 bound RBD after local refinement. The long CDRH3 loop around the RBM loop is indicated by an arrow, and the glycan linked to N343 is marked with red sphere. b Top view of 14B1 bound RBD. For simplicity, only the regions with the red dashed box in (a) are shown. The CDR loops are labeled, and hACE2 binding motifs (RBM) are highlighted in light gray on the RBD surface colored in beige. c Interactions between RBD and the CDRL2, CDRH1, and CDRH2 loops. The residues bound with CDRL2, CDRH1 and CDRH2 are shown as green stick. The color of CDR loops is similar to (b) and the residues bound with RBD are shown as gray stick. The yellow dashes represent hydrogen bonds and salt bridges. d Stereo pairs showing interactions between CDRH3 and RBD. The color scheme is similar to (c)
The 14B1 paratope buries a surface area of ~1055 Å2 on one up-RBD, calculated by PISA,26 and the heavy chain accounts for ~80%. The complementarity-determining regions of the heavy chain (CDRHs) straddle the “RBD ridge”4 (Fig. 6b) to dominate the 14B1-RBD interaction. In contrast, the light chain of the antibody shows less contact with the RBD, and mainly interact with RBD through one hydrogen bonds formed between Thr56 and Ser477 (Fig. 6c). CDRH1 and CDRH2 bind to an epitope on the RBD that faces the threefold axis of the spike, which is inaccessible in the RBD-down conformation. Asp31 from CDRH1 and Asp53 from CDRH2 form hydrogen bonds with Lys417 and Tyr505, respectively (Fig. 6c and Supplementary Table 2). Notably, the 26-residue-long CDRH3 loop (Supplementary Fig. 7) kinks at Ala93 and Ile102 to form a hook-like loop span the saddle (β5-β6) of RBD and plays a crucial role in the Fab-RBD association, accounting for 74% (645.6 Å2) of the heavy chain paratope. CDRH3 comprises a series of aromatic and hydrophobic residues at the center of the paratope and forms multiple hydrophobic interactions and hydrogen bonds with RBD, including hydrogen contacts formed between Phe98, Tyr99, Asn100, Pro100A, Arg100C, and Tyr100E of CDRH3 and Gln493, Phe490, Ser494, Try351, and Glu484 of RBD, respectively (Fig. 6d and Supplementary Table 2).
The Cryo-EM structure of 7B3 in complex with SARS-CoV-2 WT spike
We also determined the structures of 7B3 Fab in complex with the SARS-CoV-2 WT-Spike-6P and Omicron BA.1-Spike-6P (Omicron-Spike-6P) at an overall resolution of 3.5 Å and 3.4 Å, respectively, and found that 7B3 showed identical binding mode to both WT-Spike-6P (Fig. 7) and Omicron-Spike-6P (Supplementary Fig. 6). Due to the lack of details for the locally refined 7B3-Omicron-Spike-6P complex map (Supplementary Fig. 6e), we then use a relatively higher resolution complex structure of 7B3-WT-spike-6P for further analysis. Unlike 14B1, 7B3 Fab can bind RBD with both “up” and “down” conformations on the Spike (Supplementary Fig. 5), although the epitope of 7B3 on Spike is partially overlapped with 14B1 binding sites (Fig. 7e). The RBD conformation of some models generated in the 3D classification procedure can be identified with two “up” and one “down”, but only the “down” RBD region is sufficient for local refinement.

Cryo-EM reconstruction of 7B3-RBD complex and structure superposition. a (Left)The overall structure of 7B3 Fab in complex with SARS-CoV-2 S-6P trimer. One Fab (light chain in teal, heavy chain in pink) bounded RBD is in the “down” position, and the Fab bound spike protomer colored in beige and the other two in gray, and the local refinement region is boxed with black dash. (Right) Side view of 14B1 bound to RBD model after local refinement, and the N343 linked glycan is shown as red sphere. b Top view of 7B3 bound RBD. The CDR loops are labeled, RBD is shown as beige surface, and hACE2 binding sites are colored in light gray. c Interactions between RBD and CDRL3 loop. The residues bound with CDRL3 are shown in green stick. CDRL3 are in teal and the residues bound with RBD are shown in gray stick models. The yellow dashes represent hydrogen bonds and salt bridges. Residues 483–490 (VEGFNCYF) are colored in coral. d Stereo pairs showing interactions between heavy chain and RBD. The color scheme is similar to (c). e Structure superposition of 14B1-RBD and 7B3-RBD. (Left) 14B1 (slate blue ribbon model) and 7B3 (pink ribbon model) both bind to the top face of RBD. (Right) Top view of the epitopes of 14B1 and 7B3, and the epitopes of two Fabs are partially overlapped. Epitopes unique to the two antibodies are labeled separately. f Mutated residues contacted by 14B1 or 7B3 on SARS-CoV-2 WT RBD in comparison with Omicron variants are shown as spheres. Color scheme is similar to (e)
Local refinement was performed and improved the local resolution of the 7B3-RBD interface to 3.9 Å (Supplementary Fig. 5). 7B3 bind at the straddle (β5–β6) and is closely associated with the RBM loop (Fig. 7a). The total buried surface area is 1023 Å2 from the RBD, and the paratope constituted all six CDRs, with the heavy chain and light chain contributing 73 and 27% of the binding surface area, respectively (Fig. 7b and Supplementary Table 3). The medium-resolution complex structure revealed that residues 483–490 (VEGFNCYF) located at the RBM loop predominantly contribute to the 7B3-RBD interaction by forming extensive hydrogen bonds made by residues Glu484, Asn487, Tyr489, Phe490 with Tyr47, Tyr59 in or adjacent to CDRH2, Asn92, Asn93 in CDRL3 and Pro99 in CDRH3 of 7B3, respectively (Fig. 7c, d). The interaction of 7B3 to RBD is mediated by eight hydrogen bonds and one salt bridge formed between Glu484 in RBD and Arg50 in CDRH2 of the heavy chain, in addition to van der Waals contacts. By superposing the RBD region of antibodies-RBD models, 7B3 would clash with 14B1 (Fig. 7e), suggesting that the two mAbs bind competitively, which is consistent with the competition BLI results.
The molecular basis of SARS-CoV-2 variants neutralization by 14B1 and 7B3
Although 7B3 partially affects 14B1’s interaction with RBD by competition BLI, the complex structures showed that 7B3’s epitope is not completely overlapped with 14B1’s binding footprint, and the proximal contacts made by 14B1 and 7B3 are distinct (Fig. 7e). Structural analysis showed that 14B1 recognized the peak of the RBM and sterically blocked the hACE2 binding to the RBD. Heavy-chain complementarity-determining regions (CDRs) contribute the majority of interactions with RBD, comprising ten hydrogen bonds targeting several residues between 484–505 and the residues Y351 and K417 of RBD. 14B1 efficiently neutralized WT as well as Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2), and Eta (B.1.525) but exhibited diminished inhibitory activity against most Omicron subvariants. Among 17 interface residues, Eta (B.1.525) carrying E484K mutation, Beta (B.1.351) and Gamma (P.1) carrying K417N/T and E484K mutations, and Delta carrying L452R/T478K mutations, while L452 and T478 only contribute 1 and 2 van der Waals contacts to bind to S100B (CDRH3) and H49 (CDRH2), respectively. Although K417 and E484 engaged in hydrogen bonds interaction with the heavy chain of 14B1, these two mutations slightly affect the binding capacity of 14B1 to the RBD (Supplementary Fig. 2), indicating that these two amino acids play minor roles in the binding and neutralization of 14B1.
There are more than 15 mutations in the RBD of the Omicron variants compared to the ancestral SARS-CoV-2 strain, 9 and 5 of which reside in the interface of 14B1 and 7B3, respectively (Fig. 7f). Based on amino acids sequence alignment and structural analysis, we found that K417, Y505, and Q493 of RBD formed hydron bonds with CDRH3 of 14B1 and were involved in extensive interaction between 14B1 and WT-Spike-6P, the substitution of K417N, Y505H, and Q493R in most of Omicron VOAs profoundly disrupted or reduced the binding of Omicron-Spike with 14B1 (Supplementary Fig. 9), which likely were responsible for the 14B1 neutralization resistance against most Omicron variants. Among the three contacts, residues 493 possibly play the most dominant role as Omicron variant BA.2.75 harboring Q on residue 493 became sensitive to 14B1 neutralization, and only this residue difference on the interface were observed between Omicron variant BA.2.75 and resistant variants Omicron BA.1, BA.2 and BA.3.
7B3 predominantly interacts with SARS-CoV-2 WT-spike at regions 450–457,470–472, and 483–494 using both heavy and light chains, and the contacts made by 7B3 on WT exhibited 100 and 94% amino acid sequence identity with Alpha (B.1.1.7) and Delta (B.1.617.2), respectively, which correspond to its neutralization breadth. Interestingly, 7B3 also potentially inhibits highly mutated Omicrons, including Deltacron, Omicron BA.1, BA.2, BA.3, and BA2.75, and shows weak or little neutralizing activity against other Omicron variants. The mutant residues of E484A in Omicron BA2.75 and E484A combined with Q493R in Omicron BA.1, BA.2, and BA.3 are located in the interface between the 7B3 and RBD. Although Omicron BA.1-RBD carrying E484A and Q493R showed reduced binding affinity to 7B3 than WT-RBD, it still could bind to 7B3 very well with a kinetic-derived binding affinity of 14.22 nM (Supplementary Fig. 2). Structurally, Q493 makes slight contact with 7B3, and E484 forms two hydrogen bonds and several van der Waals contacts with heavy and light chains of 7B3 (Supplementary Tables 2 and 3). It looks like E484 was essential for its neutralization activity as 7B3 lost the inhibition activity against Beta and Gamma variants, and only one mutation (E484K) was present in the interface of 7B3 with Beta- and Gamma-Spike. However, the E to A mutation in residues of 484 in Omicron BA.1, BA.2, and BA.3 was adorable and still sensitive to 7B3’s neutralization.

{kind=link}